Garlic and allopurinol alleviate the apoptotic pathway in rats’ brain following exposure to fipronil insecticide
Article information

Abstract
Fipronil can cause oxidative tissue damage and apoptosis. Our goal is to evaluate the antiapoptotic impact of garlic or allopurinol against fipronil neurotoxicity. Thirty-six mature male albino rats were separated into control, garlic aqueous extract (500 mg/kg), allopurinol (150 mg/L in their drinking water), fipronil (13.277 mg/kg), garlic+fipronil, and allopurinol+fipronil. Our results revealed that fipronil induced a significant increase in brain malondialdehyde, protein carbonyl levels as well as enzymatic antioxidant activities (superoxide dismutase, catalase, glutathione peroxidase, and xanthine oxidase), but glutathione-S-transferase recorded a significant decrease as compared to the control. In addition, fipronil significantly up-regulated the brain pro-apoptotic (Bax) and caspase-3 mRNA gene expression and induced DNA fragmentation but caused down-regulation in anti-apoptotic (Bcl-2) mRNA genes expression. Interestingly, co-administration with garlic or allopurinol improved the lipid peroxidation, antioxidant disturbance, and apoptosis induced by fipronil in the brain tissues. In conclusion, garlic or allopurinol reduced fipronil-induced apoptosis and reduced oxidative tissue damage, most likely through enhancing the tissue antioxidant defense system.
Introduction
Fipronil (FPN) is a second-generation N-phenylpyrazole insecticide. It can block the gamma-aminobutyric acid (GABA)-regulated chloride channels which cause depression in the central nervous system (CNS) and pests’ death [1]. Its selective toxicity to invertebrates compared to mammals is due to the difference in binding between mammalian and insect GABA receptors [2]. Moreover, FPN may be metabolized into numerous metabolites such as FPN sulfone, which has a greater affinity to mammals’ channels than insects’ channels causing harmful effects on the non-target organisms, including humans [3].
Many researchers have stated that the toxic effects of FPN in non-target organs are due to an increase in reactive oxygen species (ROS) production, which consequently causes tissue damage [4] and apoptosis [5]. ROS can form lipid peroxyl radicals and hydroperoxides when they target lipids with carbon-carbon double bonds, mainly polyunsaturated fatty acids [6,7], resulting in cell damage after the elevation in the levels of malondialdehyde (MDA) [7] and protein carbonyl (PC). This elevation can disrupt the antioxidant enzymes including, superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx). Additionally, it disturbs the oxidative phosphorylation of mitochondria, resulting in adenosine triphosphate exhaustion [8], which consequently leads to pro-apoptotic protein (Bax) overproduction, and anti-apoptotic protein (Bcl-2) downregulation [9], and increasing production of caspase-3 leading to apoptosis [10].
The usage of antioxidants to protect from oxidative injury induced by pesticides has recently developed as an important way to hinder the construction of ROS [11]. Garlic, Allium sativum, is conventionally used for both culinary and medicinal purposes [12]. The beneficial uses of garlic on health problems could be credited to its phytochemical components. The lipophilic, as well as hydrophilic organosulfur compounds in garlic, have been confirmed to protect against oxidative injury by activating antioxidant enzymes [5]. Therefore, the protective role of garlic extract to the toxic effects is due to microsomal lipid peroxidation inhibition, antioxidant/detoxification system improvement, and the free radical scavenging effect [13]. Furthermore, allicin is a potent natural antioxidant obtained from garlic [14] and reported as ameliorating factor for many known pesticides and xenobiotics.
Allopurinol (ALP) is considered a competitive inhibitor of xanthine oxidase (XO) and used in the treatment of gout. It is an efficiently used drug in the therapy of certain central nervous system diseases [15]. This property may be attributed to the modulation of superoxide dismutase activity [16] or the increased adenosine level after XO activity inhibition [17]. Moreover, in experimental models of focal ischemic stroke, ALP pretreatment provided neuroprotection [18] by reducing the ROS production [19] and reactivating the mitochondria to produce more adenosine triphosphate. As it is a XO inhibitor, it decreases the production of hypoxanthine and prevents excessive adenosine triphosphate depletion [20].
Although the brain has different components that achieve a variety of important functions, there is little known about the effects of FPN on this vital organ. Therefore, we have looked into the ability of FPN to cause oxidative stress as an underlying mechanism of apoptosis initiation to estimate the neuroprotective effect of garlic or ALP. The lipid peroxidation occurrence (MDA), oxidant/antioxidant enzyme activities (SOD, CAT, GST, GPx, and XO), and protein carbonyl levels were first evaluated for this purpose. Subsequently, we have assessed the expression of apoptotic genes expression, histopathological alterations of the brain, and DNA fragmentation as an end product of the apoptosis process.
Materials and Methods
The pesticide and antioxidants used
A commercial product of FPN (Ptera Nil 2.5% emulsifiable concentrate) was provided by Suez Canal Trade & Agricultural Development (Egypt). The garlic aqueous extract was prepared by dissolving 25 g of garlic bulbs (Ministry of Agriculture market, Egypt) in 50 mL of distilled water. Rats were given 500 mg/kg body weight of fresh garlic extract orally using a stomach tube attached to a graduated syringe [21]. GlaxoSmithKline S.A.E. provided a commercial ALP product under license from Aspen Pharma Trading Limited (Ireland). It was produced in drinking water regularly at a dose of 150 mg/L [22]. The salts were obtained from MP chemical company (France) and Sigma Chemical Company (USA) was used as a provider for all chemicals and substrates.
Experimental procedure
Animals
In this investigation, we utilized 36 mature male Wister albino rats (Egyptian Organization of Biological Products and Vaccines, Egypt) with a mean weight of 160±10 g at the start of the trial. The animals were housed for two weeks in a random order (Mammalian Toxicology Department-Central Agricultural Pesticides Laboratory-Agriculture Research Center, Egypt) for adaptation. They were monitored daily for any abnormal symptoms. To ensure hygienic conditions, the animals were confined in plastic cages with grilled stainless-steel lids holding a layer of sawdust that was replaced every three days. They have nourished well-balanced food and access to free tap water. The cages were placed in an air-conditioned room with a temperature of 23±2 °C, relative humidity of 55% (55–70%), and a regular light/dark cycle.
Experimental groups
Animals were divided into six experimental groups (n=6). Animals in the control group (C) were given distilled water (1 mL/animal) by gastric intubation every day for 28 days. Rats orally received a dose of (500 mg/kg) aqueous garlic extract in the garlic group (G) daily all over the experimental period [21]. Rats were administered (150 mg/L) ALP dissolved in their drinking water [22] all over the experimental period in the allopurinol group (A group). In the FPN group (F), rats orally received (13.277 mg/kg equivalent to 1/10 LD50) FPN daily all over the experimental period [5]. In the garlic and FPN group (G+F), rats orally received equal doses of garlic extract as in the G group two hours before FPN administration (13.277 mg/kg). In the ALP and FPN group (A+F), rats orally received the same doses of ALP as in the A group two hours before FPN administration (13.277 mg/kg).
Specimens
Animals were lightly anesthetized after 28 days and brain tissues were immediately removed from sacrificed animals, washed with saline, dried, and kept frozen at −80 °C for biochemical parameters assay. For histopathological examination, part of these tissues was kept in formaldehyde (10%).
Preparation of the brain tissues
The brain tissues were homogenized (1:10 w/v) in precooled 50 mM potassium phosphate buffer, pH 7.5, with 1 mM EDTA. The homogenate was centrifuged at 10,000×g for 15 minutes at 4 °C in a Sigma Laboratory Centrifuge (3K30, Germany), and the supernatant was collected and kept at −80 °C. Bradford’s colorimetric technique was used to determine the protein content [23] using bovine serum albumin as standard.
Biochemical analysis
The biomarker for lipid peroxidation, MDA, was measured in brain tissues using thiobarbituric acid [24]. The activity of SOD was measured using pyrogallol as in the procedure of Marklund and Marklund [25], while CAT activity was measured using hydrogen peroxide as in the method of Aebi [26], and GPx activity was measured using the reduced glutathione as in the technique of Necheles et al. [27]. The GST activity was assessed using 1-chloro-2, 4-dinitrobezene, and reduced glutathione according to the method of Habig et al. [28]. According to the Hatano et al. [29] method, the XO activity was detected in 50 mM potassium phosphate buffer using xanthine as substrate. The protein carbonyl level was determined based on the method of Levine et al. [30]. In this method, the carbonyl groups react with 2,4-dinitrophenylhydrazine to form 2,4-dinitrophenylhydrazone and formed hydrazone spectrophotometrically assessed.
Assessment of apoptotic genes expression
The RNeasy Mini Kit (Qiagen, Germany) was used to extract RNA, and Revert Aid Reverse Transcriptase (Thermo Fisher) was used to generate the matching cDNA. Metabion (Germany) provided highly pure salt-free primers of Bax, Bcl-2, caspase-3, and β-actin (Table 1). A 25 μL total reaction volume [12.5 μL 2×SYBR Green (Qiagen PCR Master Mix, Germany), 1 μL primers, 2 μL cDNA, and 8.5 μL of RNase Free Water] was used for quantification. Each cycle (40 cycles) comprised 5 minutes of denaturing at 94 °C, 30 seconds of annealing at the proper annealing temperature, and 30 seconds of polymerization at 72 °C. After amplification, the dissociation stage was used to confirm the specificity of the products. The Stratagene MX3005P software was used to do the quantitative analysis, and changes in the mRNA gene expression were assessed using the “ΔΔCt” protocol [31].

Oligonucleotide primers used in SYBR Green RT-PCR.
Assessment of the quantity of DNA fragmentation
The diphenylamine assay was used to quantify the DNA fragmentation in the brain tissue [32].
Examination of histopathology
Brain specimens were embedded in paraffin for the preparation of 4 microns thickness paraffin beeswax tissue blocks. For examination using a light electric microscope, the obtained brain tissue sections were collected on glass slides, deparaffinized, and stained with hematoxylin and eosin stain [33].
Statistical analysis
The software program GraphPad Prism version 7.02 (GraphPad Software, Inc. CA, USA) was used to do all statistical analyses. The data is presented as the mean±standard error (SE) for six animals in each group. After analysis of variance (ANOVA), Tukey’s multiple comparisons post hoc analysis was used to estimate the comparisons between the groups. The statistical significance level was set at p<0.05 probability.
Results
Oxidant/antioxidant status
FPN intoxication resulted in an 84.77% and 71.55% significant rise in the brain level of MDA and protein carbonyl, respectively, as compared to the control group (p<0.005). In addition, the activities of SOD, CAT, GPx, GST, and XO (Table 2) were increased by 464.94%, 262.20%, 92.16%, 44.53%, and 132.6%, respectively. In comparison to the FPN-intoxicated group, the ALP co-administered group showed significant improvements (p<0.001) in brain MDA, SOD, XO, and CAT levels, but no significant changes in protein carbonyl, GPx, and GST levels. The neurotoxicity of FPN was reversed by co-treatment with ALP, as the examined parameters exhibited non-significant changes, except for SOD, GPx, and XO activities (Table 2).

The protective role of garlic or allopurinol in the fipronil-induced alteration on brain oxidant/antioxidant status of male rats.
The brain MDA level, protein carbonyl level, GPx, SOD, CAT, and XO activities were significantly improved (p<0.001) in the garlic co-treated group compared to the FPN-intoxicated group, however, GST activity was not significantly modified. Although, garlic has normalized the levels of protein carbonyl, CAT, and GST (Table 2), the levels of MDA, SOD, GPx, and XO were still significantly raised.
Relative quantity of apoptotic genes
After FPN administration, the brain Bax (18.79-fold) and caspase-3 (95.35-fold) gene expression were significantly up-regulated (p<0.0001) when compared to the control group, whereas the relative quantity of Bcl-2 (0.049-fold) was significantly down-regulated (Figure 1). The co-treatment with garlic or ALP significantly down-regulated (p<0.0001) the apoptotic genes expression (Bax and caspase-3) and up-regulated (p<0.05) the anti-apoptotic Bcl-2 gene expression as compared to the FPN-intoxicated group (Figure 1).
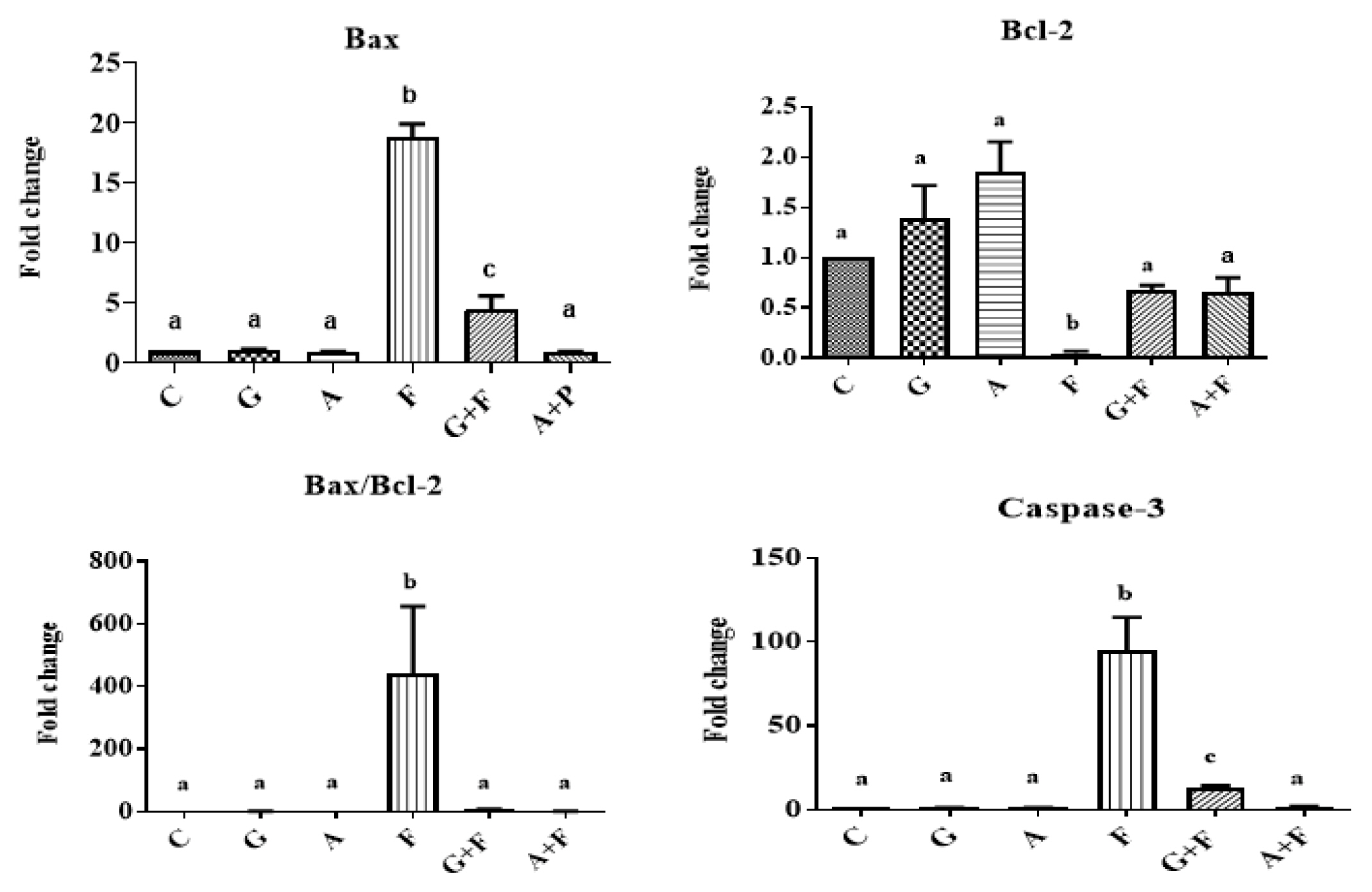
The protective role of garlic or allopurinol in the fipronil-induced dysregulations on the brain Bax, Bcl-2, Bax/Bcl-2, and caspase-3 mRNA genes expression of male rats. Each value represents the mean of six animals means±SE with different letters indicated the variations between the groups within the same column using Tukey’s honestly significant difference (p<0.05) test. Control group (C); garlic group (G); allopurinol group (A); fipronil group (F); garlic+fipronil group (G+F); and allopurinol+fipronil group (A+F).
Furthermore, co-treatment with garlic or ALP was able to normalize the expression of apoptotic genes, except for Bax mRNA (4.4-fold) in the case of garlic co-treatment, which was significantly increased (p<0.001) in comparison to the control group (Figure 1).
Quantification of DNA fragmentation percentage
When compared to the control group, the percentage of brain DNA fragmentation was significantly higher (p<0.0001) following FPN intoxication. Moreover, the garlic or ALP co-administered group showed a significant decrease (p<0.001) in the brain DNA fragmentation percentage, as compared to the control percentage. The brain DNA fragmentation caused by FPN was normalized by co-treatment with ALP, however; it was not normalized by garlic (Figure 2).
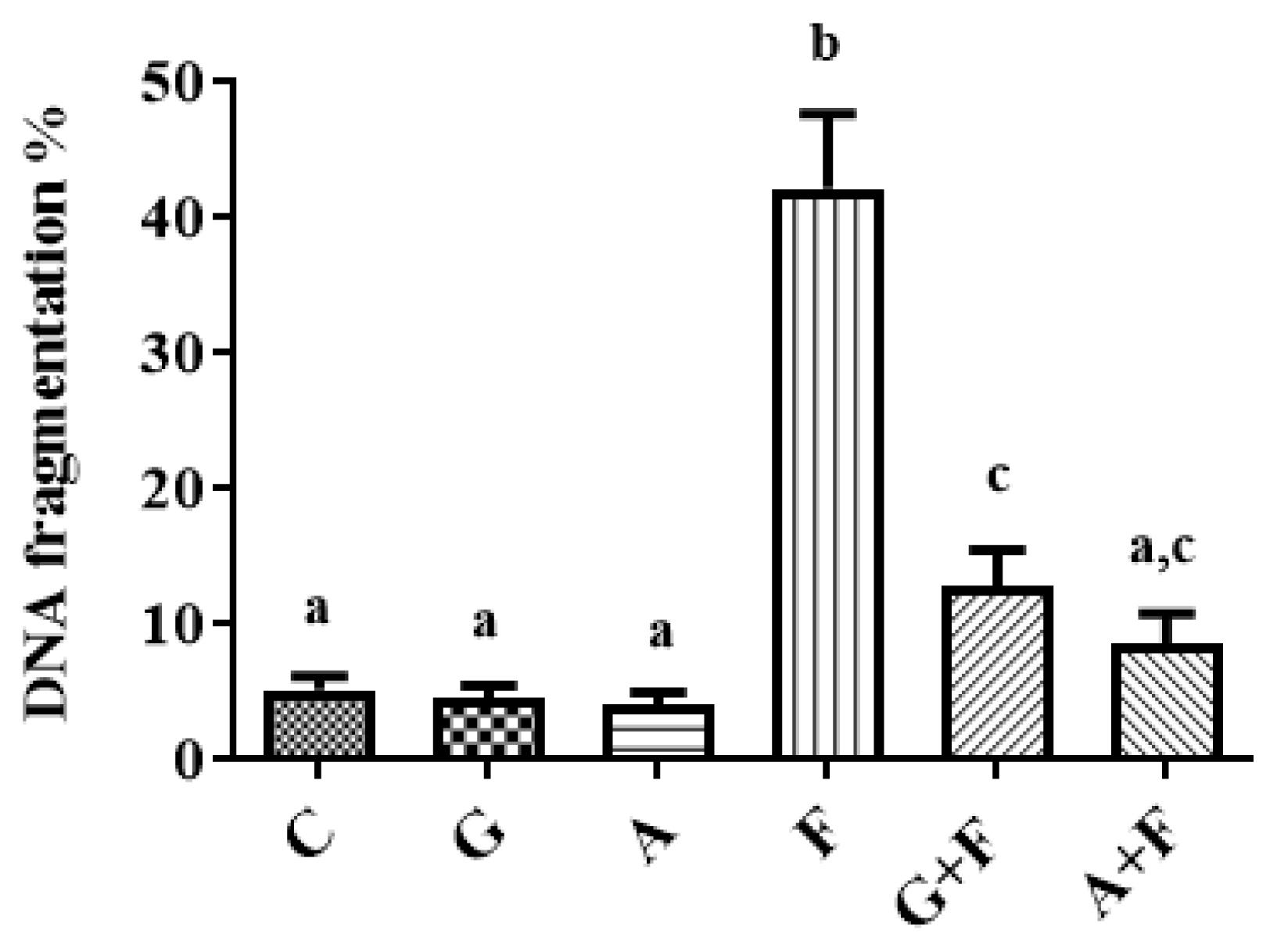
The protective role of garlic or allopurinol in the fipronil-induced DNA fragmentation on the brain of male rats. Each value represents the mean of six animals means±SE with different letters indicated the variations between the groups within the same column using Tukey’s honestly significant difference (p<0.05) test. Control group (C); garlic group (G); allopurinol group (A); fipronil group (F); garlic+fipronil group (G+F),; and allopurinol+fipronil group (A+F).
Histopathological results
Normal structure of the cerebral cortex with covering meninges, hippocampus noticed in the control (Figure 3A), garlic (Figure 3B), and ALP (Figure 3C) groups. Intracellular vacuolization was noticed in the neurons of the subiculum in the hippocampus (arrow) with congestion in the cerebellum blood vessels (star) in the FPN group (Figure 3D). Although there was no histopathological alteration in the brain section (Figure 3E) of the G+F group, focal gliosis (arrow) in the striatum was recorded in the A+F group (Figure 3F).
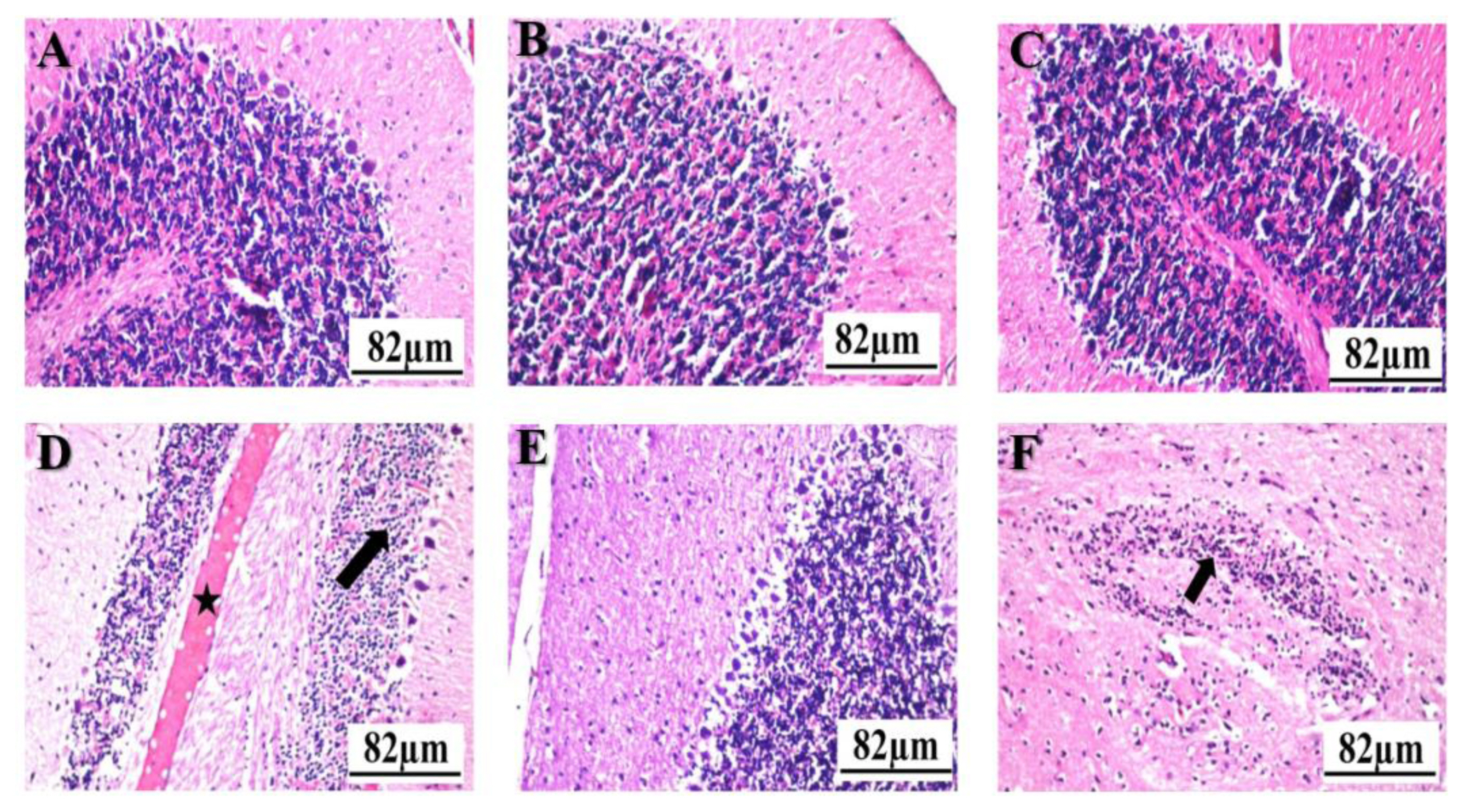
Histopathological photographs of brain sections stained with hematoxylin and eosin: The control (A); garlic (B); allopurinol (C); and garlic+fipronil (E) groups showed normal morphological assemblies with a well-arrangement appearance. Vacuolization (arrow) and congestion in the blood vessels (star) were noticed in the brain of the fipronil group (D); Focal gliosis (arrow) in the striatum was recorded in allopurinol+fipronil) group (F); Scale bar 82 μm.
Conclusions
The increased lipid peroxidation after pesticide exposure is an important indicator of the pro-oxidant stimulus [34]. In this respect, the level of MDA has been utilized as a measure of oxidative stress caused by lipid peroxidation [7]. Similarly, protein carbonyl content is the most used protein oxidation marker [35]. In the current study, the mean levels of the brain MDA and protein carbonyl were significantly increased after the administration of FPN. This increase may result from oxidative degradation of polyunsaturated fatty acids due to lipid peroxidation that disturbs the membrane structure-function [36], induces oxidative modification of membrane-bound proteins, and increases the level of protein carbonyl [37]. This hypothesis is in a line with the study of Awad et al. [38], who showed an increase in MDA and protein carbonyl levels in the brain tissues after intoxication with FPN. Additionally, FPN might elevate the brain activities of SOD, GPx, and CAT due to the protective effect of these enzymes. This defensive effect can occur by scavenging superoxide anions and hydroxyl ions, where FPN can attack the mitochondria and cause oxidative phosphorylation with over accumulation of O2• − [8]. As an adaptive response, this augmentation can increase SOD activity, which can then lead to excessive H2O2 production, resulting in CAT and GPx production, indicating the whole antioxidant defense mechanism’s inability to protect tissues from mechanical damage [39].
In the current study, the increased activity of XO in the brain tissues of the fipronil group may be due to the toxic effects of FPN on mitochondria which leads to depletion of adenosine triphosphate [40]. This diminution can result in the production of adenosine diphosphate, which promotes the creation of hypoxanthine and xanthine, resulting in improving the activity of XO. This increase can initiate an increase in superoxide radicals and hydrogen peroxide, which raises the risk of oxidative stress and apoptosis [41].
The current findings suggested that FPN can cause oxidative stress by disrupting the enzymatic and non-enzymatic defense systems, as well as trigger apoptosis via an intrinsic pathway, including an increase in the apoptotic (Bax) gene and a reduction in the anti-apoptotic (Bcl-2) one. Accordingly, the high expression of Bax mRNA, after FPN administration, can induce apoptosis by interacting with Bcl-2, which permeabilizes the mitochondrial membrane and disrupts transmembrane potential [42]. This impact can cause mitochondrial cytochrome-c to leak into the cytoplasm, resulting in the creation of an apoptosome complex. This complex can activate caspase-9, which sequentially activates caspase-3 to initiate apoptosis by releasing caspase-activated deoxyribonuclease, which fragments DNA [5].
The histopathological changes in the current study may identify the oxidant/antioxidant imbalance and apoptosis induction where intracellular vacuolization in the neurons with congestion in the blood vessels was noticed in the brain after FPN administration [43]. These shifts may be due to the FPN-brought reactive oxygen species, which results in the damage of cell membrane components [44]. Moreover, stimulation of a significant inflammatory response due to the release of constituents of the cells to the extracellular environment after plasma membrane rupture can cause inflammation that leads to tissue remodeling with fibrosis [45].
Now, there is an enlarged use of natural products as an antioxidant instead of synthetic chemicals [46]. In this respective, the alleviation efficacy of garlic may be due to its antioxidant properties as the garlic can scavenge reactive oxygen species [47], specifically superoxide radicals [48], and neutralize hydrogen peroxide, by causing the increase in the activity of SOD, CAT, and GPx, leading to prevention of hydrogen peroxide-induced cell damage [49]. Garlic also can obstruct lipid peroxidation by scavenging the hydroxyl and peroxyl radicals [50]. Moreover, garlic can hinder the oxidation of low-density lipoproteins, DNA oxidative damage, and glutathione oxidation in endothelial cells [50] that diminishes the ROS concentrations during ischemia, and attenuates mitochondrial dysfunction [51]. Subsequently, garlic can prevent the FPN-induced oxidative damage which inhibits the release of cytochrome-c, prevent the activation of caspase-3, and lastly leads to apoptosis prevention and DNA fragmentation inhibition [52].
Remarkably, the protective efficacy of ALP against neurotoxicity of FPN may have resulted as it’s a competitive inhibitor of XO or due to its ability to scavenge free radicals [53]. In this issue, it was stated that ALP has a protective role against oxidative stress by inhibiting the creation of ROS [54], mitochondrial swelling [55], and lipid peroxidation [55]. It also improved the resynthesis of adenosine triphosphate during reperfusion [55], protected against necrosis and apoptosis [56]. Moreover, ALP could significantly decrease the infarct volume and inhibit nitrosative stress and fibrosis [57] by reducing the activation of nuclear factor-kappa B [58]. Therefore, our findings assumed that ALP could protect from the brain apoptotic pathway of FPN via its inhibition the activity of XO which may down-regulate the levels of the apoptotic genes (Bax and caspase-3) and block the mitochondrial pathway of apoptosis.
In our study, garlic or ALP co-treated groups also relieved the histopathological changes where garlic pre-treatment resulted in no histopathological alterations, but ALP co-treated group showed focal gliosis in the striatum. In this respect, garlic or ALP may reduce the indications of acute inflammation on histology in the FPN-treated animals as garlic has antioxidant [59] and anti-inflammatory properties [60]. The anti-inflammatory activity of garlic may occur via the suppression of pro-inflammatory cytokines expression [61]. Moreover, ALP has an inhibitory efficiency against inflammatory response by reducing the production of free radicals [62] and inflammatory cytokines [63].
In conclusion, this study verified that oxidative injury has a vital role in the FPN-induced brain apoptotic pathway; however, garlic or ALP has prevented this pathway. We assumed that the garlic or ALP could have a probable antioxidant activity against the neurotoxicity of FPN. This impact may occur by boosting the antioxidant defense mechanism of the cell which inhibits the expression of apoptotic genes (Bax and caspase-3) and up-regulates anti-apoptotic gene expression (Bcl-2), leading to the suppression of apoptosis and DNA fragmentation.
Notes
Ethics Approval
The study protocols were approved by Institutional Animal Care and Use Committee-Cairo University (CU/I/F/60/18).
Consent to Participate
This is an experimental animal study.
Consent for Publication
This is an experimental animal study.
Conflict of Interest
The authors declare no competing interests.
CRediT author statement AA: Resources, Methodology, Data curation, Formal analysis, Software, Investigation, Validation, Visualization, Writing-Original draft; MA: Data curation, Formal analysis, Software, Investigation, Validation, Visualization; KI: Conceptualization, Project administration, Resources, Methodology, Data curation, Formal analysis, Software, Investigation, Validation, Visualization, Writing-Review & Editing.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors