Introduction
Heavy metals such as mercury, lead, and nickel accumulate in air, water, and land due to increased industries resulting from development [1]. Mercury (Hg) exposure has increased globally due to industrial activities, gold mining, medicinal uses, and fossil fuel combustion [2]. Occupational mercury chloride (HgCl2) exposure resulted in neurotoxicity, including cognitive impairment, motor dysfunction, central auditory system dysfunction, nephrotoxicity, hepatotoxicity, and mortality [3]. It primarily damage brain tissue, causing focal brain damage that can affect the function of brain parts like the cerebellum, hippocampus, and visual cortex and cause cognitive and behavioral changes [4]. Many investigations have shown that mercury causes toxicity by producing reactive oxygen species (ROS) and rapidly depleting beneficial antioxidant enzymes such as catalase, superoxide dismutase (SOD), and glutathione peroxidase (GSP). ROS promotes mitochondrial malfunction, neuroinflammation, and apoptosis, all of which contribute to neurodegenerative diseases and neuronal cell death [5]. Mercury accumulation in brain tissue increases the expression and reactivity of N-methyl-D-aspartate (NMDA) receptors. Activation of NMDA type glutamate receptors enhances Ca2+ entry into neurons, which motivates neuronal cell death pathways. Furthermore, Ca2+ promotes ROS production via the mitochondrial pathway [6,7].
The hippocampus is responsible for integrating two forms of memory: event storage and recall. The biochemical activities proceed along neuronal transmission channels, with the hippocampus playing a critical role in cognitive and behavioral analyses [8]. Previous investigations in the hippocampus confirmed the existence of elevated amounts of Hg in the brain parenchyma following prolonged exposure to HgCl2 [9] and compromised its crucial role in learning and memory [10]. Despite HgCl2’s limited capacity to penetrate biological barriers, it can nevertheless result in motor impairments because it induces apoptosis and reduces the number of neurons and astrocytes in the motor cortex [11].
Despite drug development research, treatment for neurological disorders remains abysmal due to modern drugs’ failure to regenerate central nervous tissues and their adverse side effects. Because oxidative stress is a common occurrence in metal toxicity, it is critical to comprehend the interaction between antioxidants and neurotoxicants like HgCl2. The antioxidant activity of medicinal plants is primarily responsible for the benefits associated with antioxidant consumption [12].
A. paniculata is a medicinal plant with much potential for treating various diseases [13]. A. paniculata contains a variety of compounds, including labdane diterpenoid lactones, flavonoids, and other compounds. It has a diverse set of pharmacological properties that have been demonstrated [14]. In Asia, America, and Africa, the plant has been used to treat various illnesses [15]. The Indian Pharmacopoeia includes this plant and is used in approximately 26 Ayurvedic formulations [16,17]. Andrographolide is the main medicinal component of A. paniculata [18]. Antioxidant, antimicrobial, anti-inflammatory, and antihyperglycemic properties are also found in A. paniculata [19]. Andrographolides are quickly excreted from the body through the urine and digestive tract. A. paniculata is an ideal substance for treating and preventing many ailments due to its wide distribution to the organs and regulatory actions [20]. Antioxidant-rich herbal plants protect against oxidative stress-related diseases such as neurodegenerative disease, cancer, inflammation, and cardiovascular disease [21]. This study examines whether A. paniculata could protect rats from HgCl2-induced memory loss by assessing acetylcholinesterase, neurotransmitter concentrations, and oxidative stress. In addition, the study assessed the histology and histochemistry of the rats’ hippocampus and cerebellum.
Materials and Methods
Mercury chloride, (BDH Chemicals Ltd, England) was used as neuronal toxicant. Vitamin C (Ascorbic acid; 70 mg/tablet) produced by Micro Labs Limited with NAFDAC number A4-6344, was obtained from a reputable pharmaceutical store (M.U.B Pharmaceutical Enterprises Ltd.) Zaria, Kaduna state, used as standard drug for Antioxidant. Ketamin (Sigma-Aldrich Co. LLC St. Louis, USA), Phosphate Bufffer Saline (PBS), 70% Ethanol (Sigma-Aldrich Co. LLC St. Louis, USA), Distill Water, measuring cylinders and beakers.
Plants collection and authentication
A. paniculata aerial pieces were obtained from Idoani, Ose Local Government Area, Ondo State. The plant specimen was identified and certified in the herbarium division of Ahmadu Bello University’s Department of Botany in Zaria, and voucher number 1331 was issued for reference.
Extraction
The A. paniculata plants were air dried at room temperature to a consistent weight and pulverized with a pestle and mortar. 500 g of dry plant material was macerated in 5 L of distilled water for 48 hours. The aqueous extract was filtered using Whatman filter paper, and the mixture was evaporated till dry using a controlled hot water bath maintained at 60–70 °C. The dried A. paniculata seed extract was stored in a 4 °C refrigerator until use. The extract was given to the rats in the form of daily suspensions in distilled water [22].
Experimental animals
Twenty-five male and female Wistar rats were obtained from the Animal House, Department of Pharmacology, Faculty of Pharmaceutical Sciences, Ahmadu Bello University, Zaria, and fed commercial grower mash feed with free access to water. The animals were housed in clean iron cages with twice-weekly bedding changes. They were acclimatized for ten days, and the rats were cared for following internationally recognized animal welfare standards. With Approval No: ABUCAUCC/2020/22, the Ahmadu Bello University Committee on Animal Use and Care (ABUCAUC) granted ethical approval.
Experimental procedure
The rats were randomly divided into five groups (n=5). The administration of mercury chloride, ascorbic acid, and A. paniculata aqueous extract is summarized in Table 1. The dose of HgCl2 (0.5 mg/kg) used in this experiment is an equivalent dose that has been linked to organ toxicity as previously reported [23]. The HgCl2 was administered via intraperitoneal injection, whereas the A. paniculata and ascorbic acid were administered via oral gavage. The experimental animals were weighed weekly before and during the study. The treatment period lasted 29 days.
Procedure for sampling
Rats were slaughtered humanely at the end of the experiment while under ketamine anaesthesia (75 mg/kg intraperitoneally). Wistar rat brain tissues were acquired and placed on ice in a Petri plate (24,25). Brain tissue was, washed in ice-cold saline, and homogenized in Phosphate Buffer Saline (pH 7.4) using a Teflon homogenizer. The brain homogenate was centrifuged at 5,000×g to eliminate cellular debris. The supernatant of the obtained brain homogenates was stored at −20 °C until use.
Assay for biochemical parameters
The supernatants were used to estimate biochemical markers such as superoxide dismutase activity (SOD), catalase activity (CAT), malondialdehyde (MDA), acetylcholinesterase, glutamate, and dopamine.
Assay of lipid peroxidation
Malondialdehyde (MDA) concentration is one of several low molecular weight end-products of lipid hydroperoxide breakdown and is used as a lipid peroxidation indicator. The production of TBARS, as assessed by the modified technique of Niehaus and Samuelson [24] and described by Akanji et al. [25] indicates lipid peroxidation. The 150 l of serum homogenate were treated with 2 mL of TBA-TCA-HCL reagent (1:1:1 ratio) and placed in a water bath at 90 °C for 60 minutes, then cooled and centrifuged at 5000 rpm for 5 minutes, and the absorbance of the pink supernatant (TBA-Malonaldehyde complex) was measured at 535 nm. The molar extinction coefficient of 1.56×10−5 cm−1M−1 was then used to compute the amount of malonaldehyde produced.
Assay of catalase (CAT) activity
Catalase (CAT) activity was determined using Aebi’s technique [26]. 100 μL of serum was added to a test tube containing 2.80 mL of 50 mM potassium phosphate buffer (pH 7.0). To initiate the reaction, 1 mL of newly prepared 30 mM H2O2 was added to start the reaction, and the rate at which it decomposed was measured using a spectrophotometer for 5 minutes at 240 nm. The catalase activity was calculated using a molar extinction value (E) of 0.041 mM−1-cm−1. One unit was the quantity of catalase that decomposes 1 μmol of H2O2 per minute at pH 7.0.
Assay of superoxide dismutase (SOD) activity
Fridovich’s [27] technique for determining superoxide dismutase (SOD) was used. This assay is based on the ability of superoxide dismutase (SOD) to inhibit auto oxidation of adrenaline at pH 10.2. To produce a 1:10 dilution of microsomes, 0.1 mL of serum was diluted in 0.9 mL of distilled water. A 0.2 mL aliquot of the diluted microsome was mixed with 2.5 mL of 0.05 M carbonate buffer. The reaction was initiated by adding 0.3 mL of 0.3 mM adrenaline. 2.5mL of 0.05 M carbonate buffer, 0.3 mL of 0.3 mM adrenaline, and 0.2 mL of distilled water comprised the reference combination. The absorbance was measured at 480 nm for 30 seconds to 150 seconds.
Assay of glutathione peroxidase (GPx)
GPx activity was assessed using Paglia and Valentine’s [28] technique with H2O2 as the substrate. The rate of NADPH oxidation at 240 nm over the course of three minutes served as an indirect indicator of the process. When hydrogen peroxide was added to a 0.1 M Tris buffer solution with a pH of 8.0, a blank without homogenate was used as a control. The measure of enzyme activity was nmol/mg protein.
Assay of acetylcholinesterase enzymes, glutamate and dopamine
An enzyme-linked immunosorbent assay (ELISA) kit was used to assess the levels of acetylcholinesterase enzymes, glutamate and Dopamine according to the manufacturer’s instructions (ACON Labs, USA).
Histological and histochemical studies
The brain was dissected out, the hippocampus and cerebellum were carefully excised, and the tissue was post-fixed in neutral buffer formalin for 24 hours before processing for histological staining. The tissues were dewaxed in two changes of xylene for three minutes each, then hydrated for three minutes each in decreasing grades of alcohol (100%, 95%, 90%, and 70%) before being rinsed in tap water to remove excess stain. The slides were then once again rinsed with tap water after being briefly submerged in acid alcohol for distinction. Scott’s tap water was used to blue the slides for 5 minutes, after which they were counter stained with Eosin for 3 minutes. After being washed in tap water, the sections were dehydrated in successively stronger alcohols and then cleaned in xylene. After that, a mountant was used to cover-slip the parts for general histology [29].
Bielschowsky silver staining
Sections that had been deparaffinized and rehydrated were then incubated in a 20% silver nitrate solution for 25 minutes at pre-warmed (37 °C), rinsed three times with distilled water, and then submerged in 10% formalin to halt the silver reaction. Then, drop by drop, ammonium silver solution was poured onto the slides, and after washing with 10% formalin, the sections were allowed to dry until they were a deep brown color. They were then rinsed with distilled water for 5 minutes, deposited in a 5% sodium thiosulfate solution for 5 minutes, dehydrated, hyalinized, and mounted [30]. After that, the tissue samples were examined and photographed using a light microscope (Olympus CHNB107MVR, Tokyo, Japan) to look for structural abnormalities at a magnification of X 250.
Results
Effects of the extract on the weight gain of the experiment rats
Rats treated with HgCl2 significantly reduced weight gain (p<0.05) when compared to controls, but rats treated with A. paniculata and HgCl2 had a significant weight gain in a dose-dependent manner. No significant difference (p>0.05) between the rats treated with the low dose of A. paniculate and the rats treated with only HgCl2 (Table 2).
Oxidative stress biomarkers
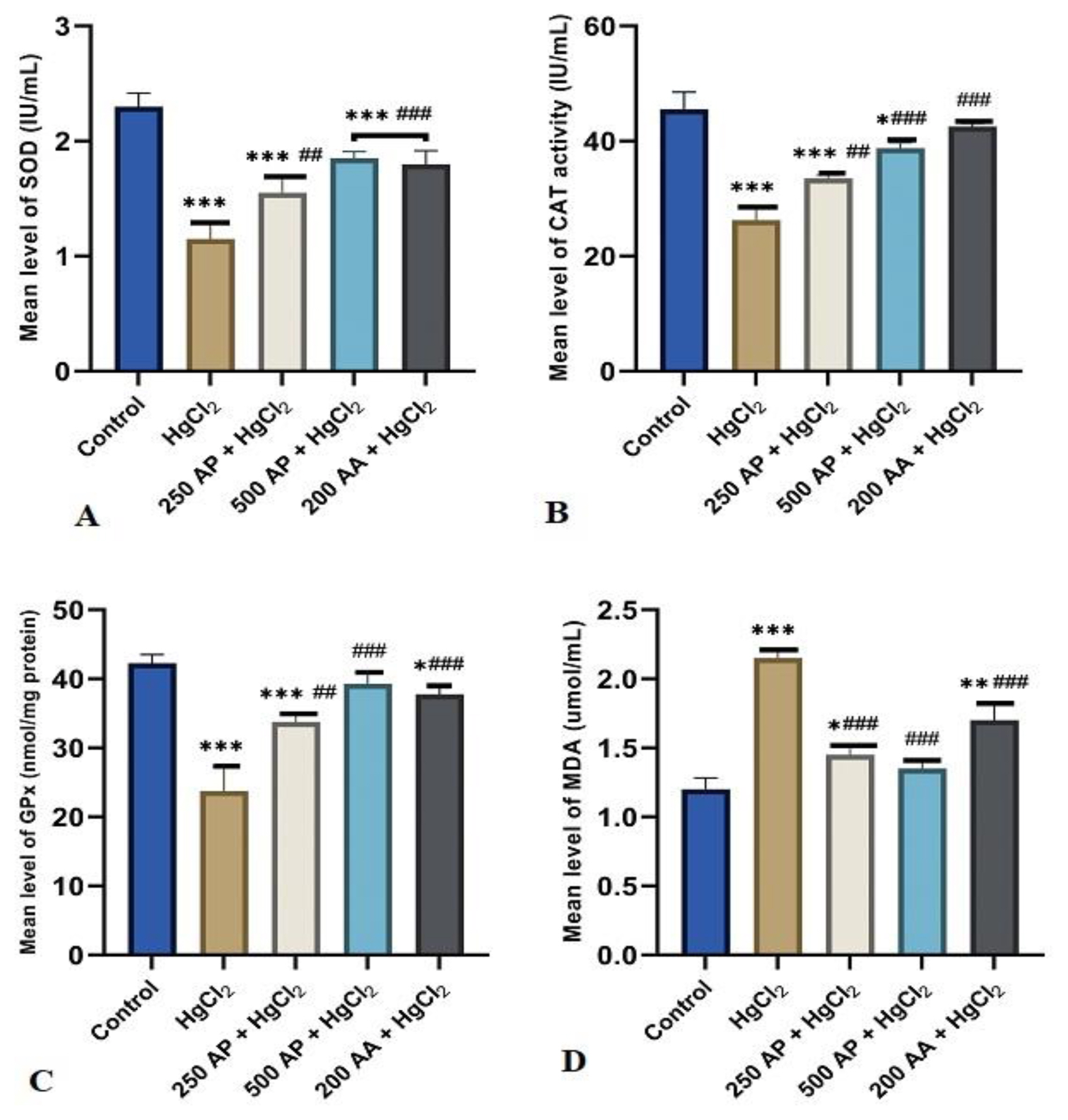
The study’s findings revealed a substantial decrease (p<0.05) in SOD, CAT, and GPx concentrations in HgCl2 treated rats as compared with control rats and rats treated with A. paniculata (250, and 500 mg/kg+HgCl2) (Figure 1A, B and C). In the case of MDA oxidative stress indicators, HgCl2 treated rats showed a significant increase (p<0.05) compared with rats treated with A. paniculate. (Figure 1D).
Neurotransmitters
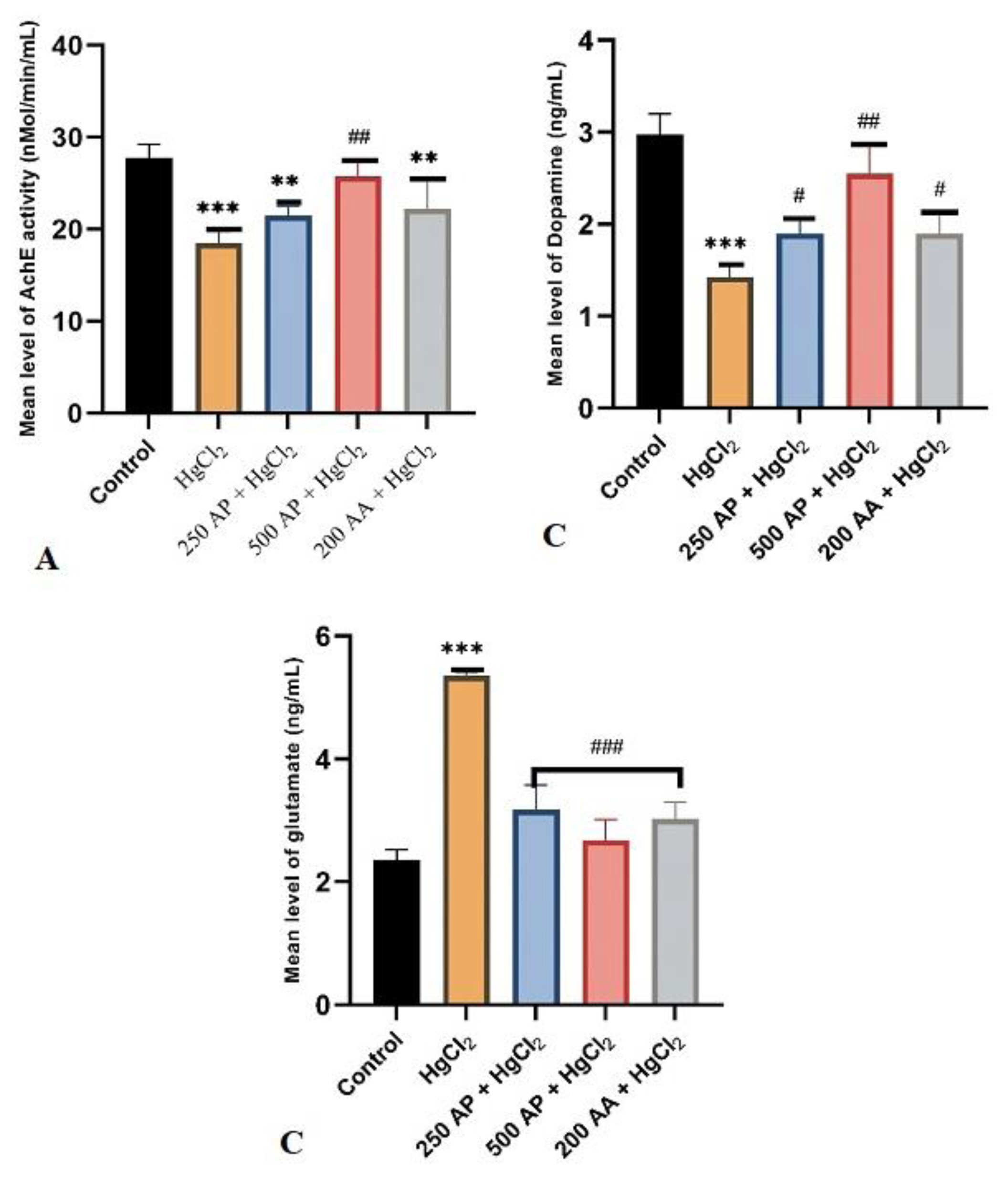
Shows the effect of mercury chloride and A. paniculata extract on Acetylcholinesterase, Dopamine, and Glutamate level. The results of the study showed a significant decrease (p≤0.05) in Acetylcholinesterase and Dopamine in HgCl2 treated rats compared with the control rats and rats treated with A. paniculata (250 and 500 mg/kg)+HgCl2, respectively (Figure 2A and B). The rats treated with A. paniculata of 500 mg/kg showed a more protective role in ameliorating the effect of HgCl2. While for the Glutamate, a significant increase (p<0.05) was observed in HgCl2 treated rats compared with the other experimental groups. No significant difference (p>0.05) was observed in all treated rat’s groups (Figure 2C).
Histological evaluation
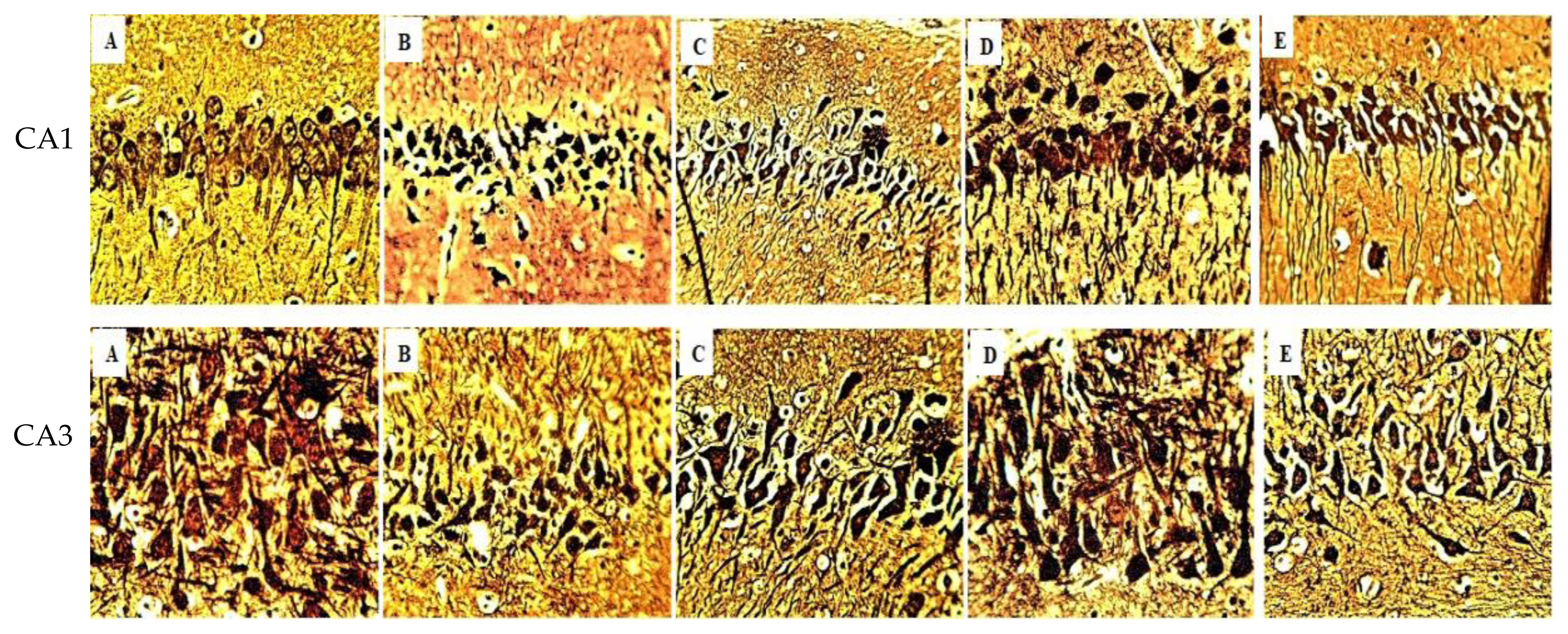
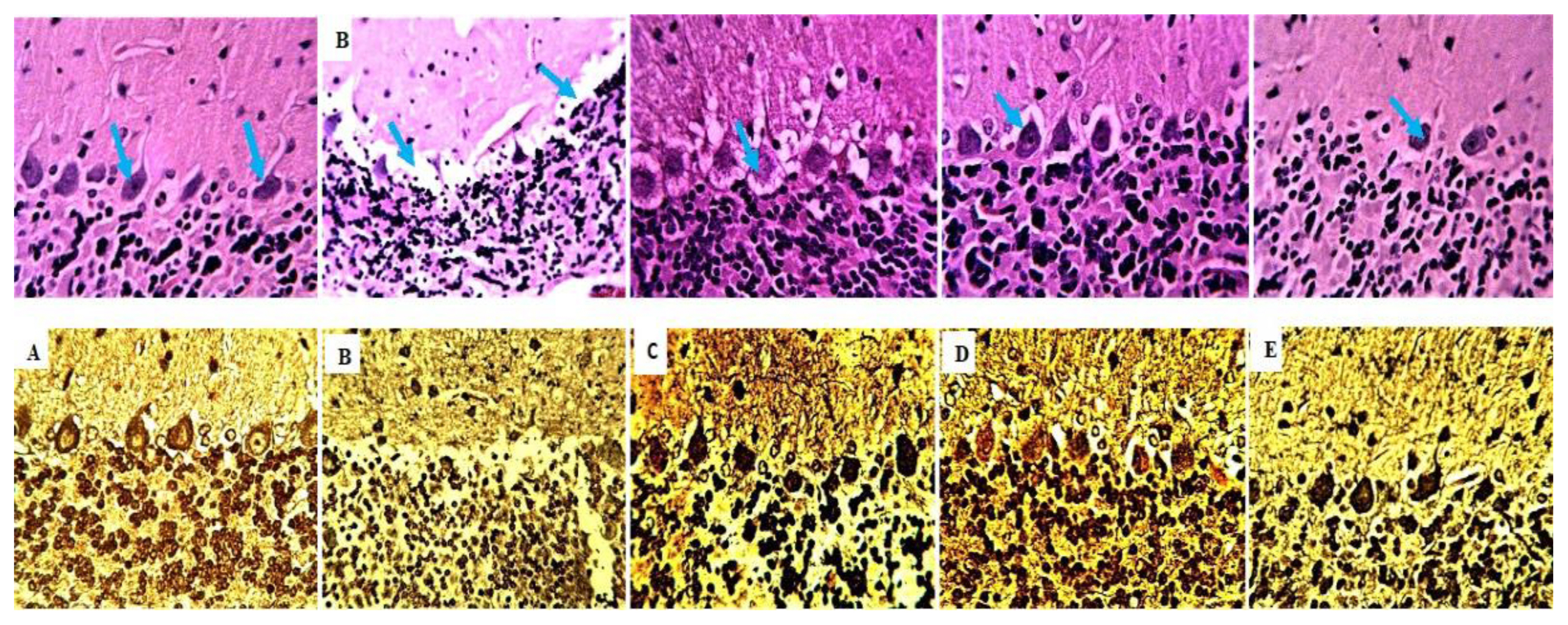
The architecture of the Hippocampus, the control’s CA1 and CA3 sections revealed densely packed, linearly oriented pyramidal cells with a well-defined form. (Figure 3A). The HgCl2-treated group, on the other hand, showed architectural deformation, particularly in the pyramidal layer, as well as cell loss and cellular disarray; the group also had cytoplasmic characteristics suggestive of cell death (Figure 3B). Compared to the control group, distortions were drastically reduced in the HgCl2+AP250 mg, HgCl2+AP 500 mg, and HgCl2+AA groups. These groups had identical cell arrangements with an intact nucleus and well-defined borders, with sporadic patches containing few, haphazardly distributed cells displaying minor reactive neurodegenerative alterations such as necrosis and certain degrees of neuronal loss (Figure 3 C, D and E). The bielschowsky stain revealed normal neuronal organization and well-organized neuronal processes (Figure 4).
In contrast, HgCl2-treated rats showed architectural deformation and neural process degeneration (Figure 4). When HgCl2+AP 250 mg, HgCl2+AD 500 mg, and HgCl2+AA treated rats were compared to the control group, mild distortion and loss of neuronal processes were seen in the CA1 area (Figure 4). In contrast, rats treated with only HgCl2 showed marked degeneration and neuronal death in the CA3. The cytoarchitecture of the molecular, granular, and purkinje cell layers seemed normal in the cerebellar control rats (Figure 5B). Rats treated with just HgCl2 exhibited a standard molecular layer with Purkinje cell layer degeneration and Purkinje cell layer degeneration, whereas the granular layer showed aberrant packed granular cells (Figure 5B). Rats given A. paniculata (250 mg/kg) had severely deteriorated Purkinje cells with the Purkinje cells, but rats given A. paniculata (500 mg/kg) had distinct molecular and granular layers, with minor degeneration of the Purkinje cells inside the Purkinje cell layer and an average orientation of the molecular layer (Figure 5D and E).
Discussion
In this study, animals given HgCl2 had a decreased gain in body weight. This result is consistent with what other researchers have observed [31,32]. The main sign of Hg toxicity is weight loss, which has been linked to decreased food consumption and rat muscle cell damage [33,34]. A. paniculata administration in this study was able to prevent such a side effect. The extract appears to have the potential to increase appetite and protect cells from oxidative stress caused by mercury, as a result, the rats’ weight improved [35]. One of the immediate effects of HgCl2 toxicity and poisoning in living organisms might be changes in oxidative state, either through excess generation of oxidants or deficiencies in antioxidant activity [4]. The current study discovered an increase in MDA and a decrease in SOD, CAT, and GPx levels in the brain tissues of HgCl2-induced rats. Regardless of the dose used, A.paniculata treatment did, however, attenuate these changes. These findings were consistent with prior research that found A.paniculata to have antioxidant effects [36–38]. The presence of bioactive compounds such as flavonoids and phenols in A. paniculata may be responsible for the antioxidant effect. Flavonoids have antioxidant properties by scavenging free radicals, preventing lipid peroxidation and playing a neuroprotective role in these studies [39]. A. paniculata has been shown to help with antioxidant defenses in previous studies [40,41]. It works by mitigating the free radicals directly. It indirectly affects mitochondrial integrity by inhibiting pro-oxidant enzymes and activating antioxidant enzymes. It can also activate enzymatic and non-enzymatic antioxidants, primarily through the Nrf2 signaling pathway [42].
Long-term exposure to HgCl2 causes neurotoxicity [43], affecting neurotransmitter synthesis and release [44]. In this current study, HgCl2 appears to reduce DA and AChE activity. A decrease in AChE and DA has also been linked to increased lipid peroxidation [45]. Previous research in rodents and humans has found a strong link between an age-related decrease in AChE, DA and increased oxidative stress. AChE activity is reduced by free radicals and increased oxidative stress [46,47]. However, the toxic and harmful effects of HgCl2 on AChE and DA activity were prevented by A. paniculata administration. A. paniculata promotes AChE activity even with HgCl2 exposure, which could be due to its ability to inhibit Hg-induced ROS and lipid peroxidation.
In the CNS, the excitatory neurotransmitter glutamate is the most abundant and is involved in neuronal plasticity, differentiation, and transmission [48]. On the other hand, excess glutamate causes abnormal depolarization of neurons, which causes excitotoxicity and death of neuronal cells [49]. The significant increase in glutamate concentration in HgCl2-treated rats may be due to the fact that HgCl2 can inhibit glutamate uptake by astrocytes at the synaptic space while stimulating glutamate release, resulting in excessive glutamate accumulation in the CNS’s extracellular space [50]. Notably, in this study, A. paniculata modulated glutamate concentration in HgCl2-treated rats. Furthermore, A. Paniculata might be able to regulate the glutamate concentration by increasing cellular uptake of glutamate to keep extracellular glutamate levels low. A. paniculata’s antioxidant potential may help prevent depolarization of cell membranes by regulating glutamate levels and preventing excitotoxicity just like other plant antioxidant potentials, as stated in a previous study [51].
Rats treated with just HgCl2 revealed neuronal degeneration, with shrunken nuclei and vacuolated cytoplasm, and some neurons with pyknotic nuclei or karyolysis at the CA1 and CA3 areas of the hippocampus, which is consistent with previous findings [52]. The degeneration of Purkinje cells within the cerebellum of rats treated with just HgCl2 showed the complete loss of nuclear materials, demonstrating the toxicity of HgCl2 [53,54]. However, treatment with A. paniculata prevented the above-mentioned HgCl2-induced morphological aberrations in the rats. The results obtained from this study are in accordance with the results documented in previous studies on the attenuating effects of Antioxidant rich plant on heavy metal-induced neurotoxicity [53]. These suggest that A. paniculata has antioxidant properties, as seen in the present study, or a combined antioxidant and anti-inflammatory role by Andrographolide, one of the major chemical constituents of the plant A. paniculate could be attributed to these positive preventive properties [40,55].
Conclusions
This research reveals that HgCl2 (0.5 mg/kg) causes an oxidative imbalance in rat’s brain tissues, resulting in oxidative stress and pathological damage. HgCl2 produced toxicity in the hippocampus and cerebellar tissues and was effectively prevented by oral A. paniculata extract therapy at a dosage of 500 mg/kg/day. As a result, we conclude that A. paniculata extract might be effective as a potential protective/preventive agent in brain tissue damage caused by HgCl2 poisoning.