INTRODUCTION
Toxic residues following accidental oil spills are one of the most important worldwide environmental concerns. Oils are comprised of many chemical species, including persistent, toxic, and carcinogenic compounds, such as polycyclic aromatic hydrocarbons (PAHs), alkylated PAHs (alkyl-PAHs), and a small fraction of non-hydrocarbons [1,2]. The majority of research on the toxicity of compounds in oils has focused on unsubstituted 16 PAHs classified as priority pollutants by the US Environmental Protection Agency [3,4]. However, alkyl-PAHs are found in higher quantities than unsubstituted PAHs in crude oils [2,5,6] and field monitoring studies have reported that higher alkyl-PAHs concentrations were detected in oil-contaminated sites, in the sediment, soil, and surface water [7,8]. Because of the predominance of alkyl-PAHs in oils, whether alkyl-PAHs are as toxic as PAHs is an important question.
Several studies reported that alkyl-PAHs were more toxic than their non-alkylated counterparts based on organism-level responses [9-13]. Studies have suggested that the toxicity of alkyl-PAHs depends on the oxidative stress resulted from the metabolic process of cytochrome P450 (CYP) induced via the interaction of xenobiotics with the aryl hydrocarbon receptor (AHR), a mechanism similar to the toxicity pathway of PAHs [14-16]. For example, Mu et al. [14] supported that oxidative stress is an important toxicity pathway, leading to developmental toxicity by showing the increase in CYP activity and the response of the antioxidant system caused by alkyl-phenanthrenes. Fallahtafti et al. [10] proposed that the toxicity of alkyl-PAHs is not a response specific to the preferential formation of particular metabolites, such as those formed from CYP metabolism. However, studies to elucidate the toxic potential and mechanisms leading to oxidative stresses caused by alkyl-PAHs are lacking although oxidative stress is believed to be a strong inducer leading to toxic effects at the organism level (e.g., survival, behavior, and reproduction).
The main goal of the study was to compare the toxicity of Ant and alkyl-Ants on the oxidative stress of Caenorhabditis elegans under constant exposure using passive dosing. C. elegans was chosen because it is a representative soil invertebrate for (eco)toxicology whose genome has been completely sequenced [17-21]. To maintain the intended exposure concentration during the experimental period, a passive dosing method using polydimethylsiloxane (PDMS) was employed. Ant, 9-methylanthracene (9-MA) and 9,10-dimethylanthracene (9,10-DMA) were chosen as model chemicals to which the soil nematode C. elegans was exposed. As oxidative stress-related end-points, reactive oxygen species (ROS) induction, superoxide dismutase (SOD) activity, total glutathione (GSH) concentration, and lipid peroxidation were measured at the same exposure concentration in the test medium, as well as at the maximum soluble concentrations of test chemicals. Oxidative stress-related genes (daf-2, sir-2.1, daf-16, sod-1, sod-2, and sod-3) and CYP metabolism-related genes (cyp35a1-5 and cyp35c1) were also investigated to identify and compare changes in genetic responses in C. elegans exposed to Ant and alkyl-Ant.
METHODS
Organisms
The soil nematodes C. elegans were maintained on a nematode growth medium at 20°C and fed Escherichia coli strain OP50. Age-synchronized young adults (3.5 days old) were gathered according to a general nematode protocol [17,18] and 100 μL of worm condensate was used for all experiments (average tissue dry-weight: 10 mg in 100 μL).
Chemicals
Ant was purchased from Sigma-Aldrich (> 99%; CAS No. 120-12-7; St. Louis, MO, USA), and 9-MA (98%; CAS No. 779-02-2) and 9,10-DMA (98%; CAS No. 781-43-1) were purchased from Tokyo Chemical Industry Co. (Tokyo, Japan). PDMS sheets were purchased from Specialty Silicone Products, Inc. (cat. SSP-M8232; Ballston Spa, NY, USA). High-performance liquid chromatography grade methanol and n-hexane were purchased from Honeywell Burdick & Jackson (Ulsan, Korea).
Experimental Procedures
Methodological details for passive dosing
PDMS sheets were cut into rectangular sheets (11 mm×109 mm×1 mm, density 1.17 g/mL) for loading with Ant and alkyl-Ant. The custom-cut PDMS sheets were washed twice with n-hexane and methanol for 2 hours each. Then, clean PDMS sheets were stored in ACS grade methanol until use. For toxicity tests, PDMS sheets were loaded with Ant and alkyl-Ant using a loading solution (methanol:water = 6:4 [v/v]). Preliminary measurements of equilibrium partition coefficients between PDMS and the loading solution for Ant and alkyl-Ant (Table 1) were used to prepare the desired concentration in the exposure medium. After 24 hours, PDMS sheets were taken from the loading solution, air-dried for 1 hour, and placed in a 6-well plate. The test medium (0.032 M potassium chloride [KCl] and 0.051 M sodium chloride [NaCl]) and PDMS were pre-equilibrated for 24 hours at 20°C before introducing worms.
Toxicity tests at the same external aqueous concentration (TEST I)
To investigate toxic responses at the same external exposure concentration for the comparison of toxic potency, three chemicals were tested at 4.0 and 8.0 μg/L, which was below the water solubility of the most hydrophobic chemical, 9,10-DMA (Table 1). Young adults (3.5 days old) were placed in a 1-mL tube and 100 μL of worm condensate was transferred to the test medium (0.032 M KCl and 0.051 M NaCl), which had been pre-equilibrated for 24 hours with each test chemical. After pre-equilibrium, the worms were exposed for 6 and 12 hours without food.
Toxicity tests at the water solubility limit of test chemicals (TEST II)
Because toxic effects generally increase with increasing internal dose of a xenobiotic chemicals [22] and organisms are exposed to the maximum external concentration of a single chemical at its maximal toxic effects at its water solubility level, oxidative stress indicators were also tested at concentrations close to the water solubility level of the three chemicals. As described above, PDMS sheets were loaded with each chemical at the maximum water soluble concentration and at half the water solubility level, as shown in Table 1. To avoid organismal-level effects such as mortality, worms were exposed to each chemical for only 3 hours without food.
Measurement of intracellular Reactive Oxygen Species Formation
C. elegans sampled after exposure were washed twice with 500 μL of S-buffer and then incubated in 50 μM 2,7-dichlorofluoroscein diacetate (D6883, Sigma-Aldrich) solution for 30 minutes at 20˚C. Next, the worms were transferred to a 96-black-well plate to detect fluorescence intensity or a slide glass to take a fluorescence image. The level of fluorescence intensity was measured using a Hidex Sense multi-detection microplate reader (Hidex, Turku, Finland) with excitation at 485 nm and emission at 530 nm. The formation of ROS was visualized using a Leica DM IL microscope with a Leica DCF 42˚C camera (Leica, Heerbrugg, Switzerland). A 2-mm tetramisole hydrochloride solution (L9756, Sigma-Aldrich) was used to fix C. elegans to take pictures of the live worms.
Superoxide Dismutase Assay
The activity of SOD was measured using an OxiselectTM Superoxide Dismutase Activity Assay kit (cat. STA-340; Cell Biolabs, Inc., San Diego, CA, USA). Collected worms were homogenized in 400 μL of cold 1× lysis buffer (10 mM Tris, pH 7.5, 150 mM NaCl, and 0.1 mM ethylenediaminetetraacetic acid). The crude homogenate was centrifuged for 10 minutes at 12 000 rpm (4˚C) and the tissue lysate supernatant was collected and stored at –80˚C until enzyme analysis. Superoxide (O2−) were generated by the xanthine/xanthine oxidase system and then detected with chromogen solution by measuring the absorbance reading at 490 nm using a PowerWaveTMXS microplate spectrophotometer (Bio Tek, Winooski, VT, USA). SOD activity was determined as the inhibition of chromogen reduction.
Total Glutathione Assay
The level of GSH was determined using an OxiselectTM Total Glutathione Assay kit (cat. STA-312; Cell Biolabs, San Diego, CA, USA). The whole sample containing 10 mg of tissue (dry weight) was homogenized in 200 μL of ice-cold 5% metaphosphoric acid and then centrifuged at 12 000 rpm and 4˚C for 15 minutes. The collected supernatant was stored at –80˚C. In the presence of NADPH, oxidized glutathione (GSSG) is transformed to reduced GSH by GSH reductase. The chromogen reacts with the thiol group to produce a colored compound that absorbs at 405 nm with a Multi-Detection Microplate Reader (Hidex Sense). The total GSH content in samples was determined by comparison with a predetermined GSH standard curve.
Lipid Peroxidation Assay
The OxiselectTM TBARS Assay kit (cat. STA-330; Cell Biolabs) was used for the direct quantitative measurement of malondialdehyde (MDA). Tissue samples were homogenized with 300 μL of phosphate buffer saline containing 3 μL of 100× butylated hydroxytoluene. Homogenized samples were centrifuged at 12 000 rpm and 4˚C for 5 minutes. The tissue lysate supernatant was collected, reacted with thiobarbituric acid at 95˚C for 60 minutes. After all sample tubes were cooled on ice, they were centrifuged at 3000 rpm for 15 minutes. Absorbance of the solution was read at 532 nm with a PowerWaveTMXS microplate spectrophotometer (Bio Tek). MDA content in a sample was determined by comparison with the predetermined MDA standard curve.
Gene Expressions
For quantitative gene expression analysis, C. elegans were homogenized in a glass Dounce tissue grinder (Wheaton, Millville, NJ, USA), and total mRNA was extracted using a NucleoSpin RNA kit (Macherey-Nagel GmbH & Co., Düren, Germany) according to the manufacturer’s instructions. Quantitative real-time polymerase chain reaction (PCR) amplification was performed using iQ SYBR Green Supermix (Bio-Rad, Hercules, CA, USA) on an Eco Real-Time PCR System (Illumina, San Diego, CA, USA). Primers were designed according to the sequences retrieved from the C. elegans database (www.wormbase.org). Twelve primers were classified under two classes as oxidative stress-related genes and CYP subfamily genes (Table S1). Gene expression was normalized to the level of the glyceraldehyde 3-phosphate dehydrogenase gene, gdp-1.
Statistical Comparison of Oxidative Markers and Gene Expressions
A parametric t-test was conducted to compare worms exposed to each test chemical and the control at 95% and 99% confidence level for gene expression tests. A single factor analysis of variance and Duncan’s multiple range test were conducted to compare the control and all exposed groups at 95% confidence level for all oxidative stress markers.
RESULTS
Oxidative Stress-related Responses at the Same Aqueous Concentration
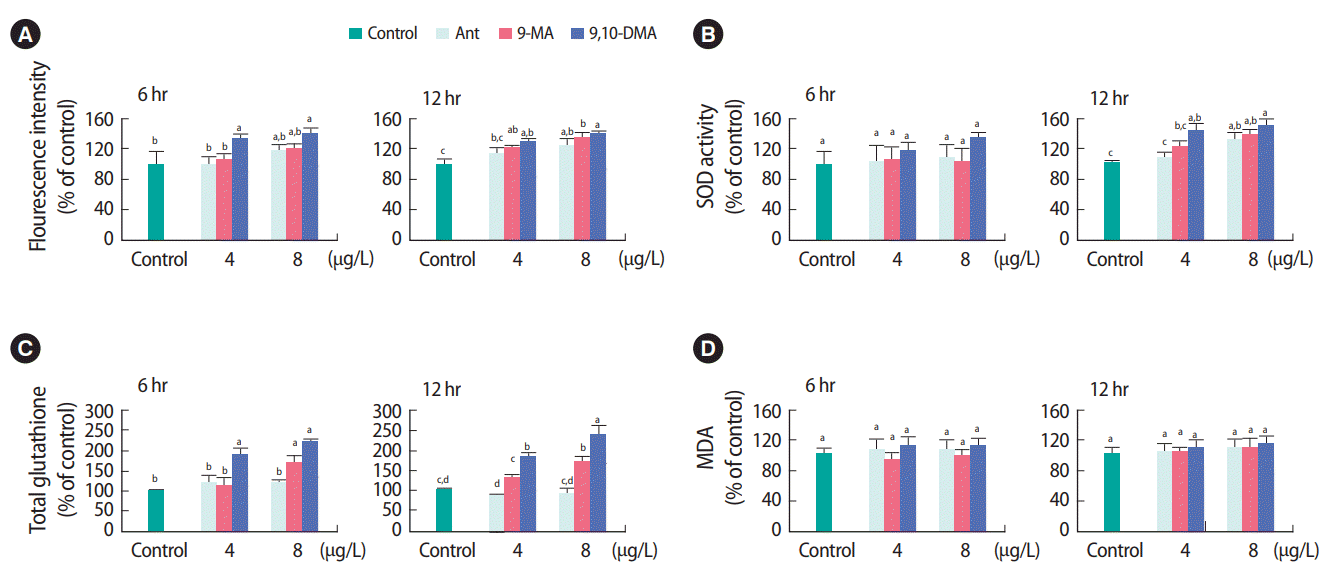
When worms were exposed to the same external concentrations, 4.0 and 8.0 μg/L of test chemicals, the formation of ROS was much higher in worms exposed to 9,10-DMA than for worms exposed to Ant and 9-MA (Figure 1A). After 6 hours of exposure, 9,10-DMA (8 μg/L) led to up to a 41% increase in ROS formation over that of the control. After 12 hours of exposure, the increase in ROS formation compared with that of the control reached 25, 34, and 40% for Ant, 9-MA, and 9,10-DMA, respectively. The activity of SOD exhibited a similar trend as that of ROS formation. When worms were exposed to 8 μg/L, 6 hours exposure to 9,10-DMA led a 35% increase in SOD activity relative to that of the control (Figure 1B). After 12 hours exposure, SOD activity in worms exposed to the maximum concentration of all three chemicals increased significantly (p < 0.05) by more than 30%. Notably, 8 μg/L 9,10-DMA produced the highest SOD activity (49% higher than that of the control) after 12 hours exposure. The level of total GSH in 9,10-DMA-exposed groups was significantly higher than those in other groups, regardless of exposure duration (Figure 1C). After 6 and 12 hours exposure to 8 μg/L 9,10-DMA, total GSH increased by 75 and 138%, respectively. No significant differences in the lipid peroxidation assay were observed among different groups (Figure 1D).
Table 2 presents the expression of oxidative stress-related genes (daf-2, sir-2.1, daf-16, sod-1, sod-2, and sod-3). In the case of exposure to Ant and 9-MA, observed gene expression was slightly greater than that of the control. However, the expression of all genes in worms exposed to 9,10-DMA was more than double that of the control at 8 μg/L after 6 hours exposure. Among the SOD genes, sod-1 gene expression increased up to 3.3-fold compared to that of the control.
Oxidative Stress-related Responses at Maximum Water Soluble Concentrations
Figure 2 describes oxidative stress-related responses obtained from TEST II using exposure concentrations at the water solubility limit of the test chemicals. The ROS levels in the presence of 9,10-DMA increased by approximately 30-37% compared with that of the control; however, the fluorescence signals caused by Ant and 9-MA exposure were not statistically different from the control (Figure 2A). In the SOD activity test, exposure to 9-MA produced a level of enzyme activity similar to that produced by 9,10-DMA, although the exposure concentration was 50 times higher than that of 9,10-DMA (Figure 2B). When worms were exposed to maximum concentrations for 3 hours, total GSH increased approximately 60% relative to that of the control, regardless of the chemical. Total GSH measured in the 1/2 water soluble concentration of 9,10-DMA (4.0 μg/L) increased by 52% relative to that of the control (Figure 2C). In the MDA assay (Figure 2D), lipid peroxidation did not show any significant response under any exposure concentration in TEST I.
As shown in Table 3, the expression of genes showed a similar trend (1.5–2-fold increases) for Ant, 9-MA, and 9,10-DMA, with the exception of abnormal dauer formation gene-16 (daf-16). The expression of the daf-16 genes reached a peak level when worms were exposed to 400 μg/L of 9-MA. After 3-hour exposure to the highest concentrations (Ant: 40 μg/L, 9-MA: 400 μg/L, 9,10-DMA: 8 μg/L), daf-16 expression was 2.60, 4.22, and 2.17 times that of the control, respectively.
Toxic Response of Xenobiotic Metabolism-related Cytochrome P450 Genes
In TEST I, only cyp35a1, among all metabolism-related genes, was significantly expressed approximately 2-fold more than in the control, regardless of the chemical (Figure 3A). When gene expression was examined at the chemical’s water solubility level, expression of all genes was increased in the order: 9,10-DMA > Ant > 9-MA (Figure 3B). In particular, expression of the cyp35a1 gene was up to 6.5-fold higher than that of the control after 3 hours exposure to 400 μg/L 9-MA. When worms were exposed to 40 μg/L Ant for 3 hours, expression of the cyp35a1 gene was 4 times that of the control.
DISCUSSION
Comparison of Toxic Potential at the Same External Aqueous Concentration
Because the free concentrations of the three chemicals in the test medium were maintained at the same level by the passive dosing method, results from TEST I can be used to compare their toxic potentials at the same external dose. It is well known that the external dose is not directly related to toxic responses at target sites that is determined by the internal dose. Since the whole-body concentration is often assumed to be a good surrogate for internal concentration [23], the differences in oxidative stress markers in C. elegans might be explained by the difference in the whole-body concentration resulting from different toxicokinetics. Reported values of the logarithm of the lipid-water partition coefficients (Klipw) for Ant, 9-MA, and 9,10-DMA are 5.28 [24], 5.52, and 5.78 [25], respectively, as shown in Table 1. According to a simple one-box bioconcentration model, the uptake rate constant is not likely to differ among test compounds, but the elimination rate constant decreases with increasing Klipw [26]. Thus, the whole body concentration of Ant, 9-MA, and 9,10-DMA could be in the order of 9,10-DMA > 9-MA > Ant at the same external concentration if other processes, such as metabolic transformation, do not differ considerably. The observed increase in oxidative stress end-points with increasing alkylation using the same external exposure concentration might be explained by the increase in Klipw, which results in higher internal concentration at the same external concentration. This hypothesis was further tested with TEST II, as discussed below.
Comparison of Toxic Potential Within the Limit of Aqueous Solubility
Figure 2A shows that the ROS level induced by exposure to 9,10-DMA was the highest, even if 9,10-DMA had the lowest external concentration. Because Klipw values for the three chemicals are within one order of magnitude, the internal concentration of 9,10-DMA would be the lowest unless there are notable differences in the metabolic transformation rates among them. In general, the expression of oxidative-stress indicators increased with increasing degree of alkylation, implying that alkylation increases the ease of electron donation to initiate a series reactions involved in the reduction of oxygen.
Most ROS are formed in the intermediate steps of the reduction of molecular oxygen (O2). The O2 in the body requires four electrons for complete reduction to water, and a one-electron step results in the formation of the O2−. The O2 plays an important role in the formation of other ROS, such as hydrogen peroxide, the hydroxyl radical, and singlet oxygen, in living systems that use electrons provided by an electron donor [27]. Alkyl groups enrich the electron density of the π electron system of aromatic hydrocarbons and facilitate electron excitation [28]. Although the mechanisms related to the π electron transfer to oxygen to form superoxide ions are not fully understood, enriched electron density is a plausible explanation for accelerated ROS formation.
Results of antioxidant enzyme activity assays support the observation that oxidative stress is increased by the formation of ROS in worms exposed to alkyl-PAHs (Figures 1B and 1C, 2B and 2C). SOD activity and the concentration of total GSH were increased in the order 9,10-DMA > 9-MA > Ant for all exposure concentrations. SOD catalyzes the reduction of O2− to hydrogen peroxide (H2O2) and molecular O2. H2O2, in conjunction with the O2, is removed by GSH peroxidase, oxidizing the reduced GSH to form disulfide glutathione (GSSG) [29]. The increase in SOD activity and the total GSH concentration showed that methyl groups in alkylated PAH favored ROS formation and were involved in increasing oxidative stress.
Changes in mRNA Expression Induced by Anthracene and Alkyl-anthracene
The daf-2 gene, which encodes the insulin-like growth factor (IGF-1) involved in longevity and stress resistance and the sir-2.1 gene, the mammalian ortholog of which is SIRT1, are also known to modulate longevity and oxidative stress responses through regulating a downstream FOXO transcription factor, daf-16. These pathways resist oxidative stress and regulate the expression of ROS detoxification genes, such as sod-1, sod-2, and sod-3 [30-33]. In TEST I, the level of gene expression induced by 9,10-DMA was slightly higher than that induced by the other compounds, whereas no significant differences were observed between Ant and 9-MA (Tables 2 and 3). The expression of mRNA partly supports that alkyl substitution of anthracene would increase ROS formation as presented in Figures 1A and 2A.
It is well known that PAHs have strong binding affinity for the AHR, and are inducers of CYP. When worms were exposed to each maximal water-soluble concentration of the test chemicals, the expression of all cyp genes increased in the order 9-MA > Ant > 9,10-DMA (Figure 3), whereas cyp gene expressions remained at a similar level in the same exposure concentration test, showing that cyp gene expression is proportional to the external concentration. This implies that residual concentrations of chemicals in the body could be an important factor that determines gene expression induced by binding of a PAH and a receptor. In particular, differences in expression of the cyp35a1 gene induced by target chemicals was enhanced depending on exposure concentrations that were strongly induced by PAHs and PCBs, as reported by Menzel et al. [19,20]. In addition, expression of the daf-16 gene was highest when exposed to 9-MA at the water solubility. The transcription factor daf-16 promotes longevity in response to many input signals, including AMP kinase, JNK-1, SIR-2, AKT, and a kinase in the insulin/IGF-1 (daf-2) pathway, in addition to ROS [34]. 9-MA at a high external concentration (400 μg/L) may affect multiple sites in an organism, and thus many stress-related signals are continuously transferred into biological cascades, leading to the increased expression of daf-16, a comprehensive stress-related transcription factor.
Comparison of Toxicity of anthracene and Alkylanthracene With Literature Values
The observed effect levels in this study were in general lower than those reported in earlier studies for Ant and 9-MA. To our best knowledge, there have been no reports on the ecotoxicity of 9,10-DMA. Sese et al. [21] investigated the toxicity of Ant on C. elegans by measuring growth at a 48-hour median effective concentration (EC50) of 800 μg/L, in which Ant was carried by acetone as a cosolvent and the reported EC50 value exceeded its water solubility. Several studies have reported that 2-day EC50 values for Ant in Daphnia were 36-750 μg/L [9,35,36], and those of 9-MA were 130-440 μg/L [9]. The range of exposure concentration in this study was 8-40 μg/L for Ant and 8-400 μg/L for 9-MA. Although they were similar to or lower than those used in other studies, the initial responses of oxidative stress at the molecular level were well identified. Furthermore, the passive dosing method provided constant exposure by overcoming the deficiencies of the conventional dosing method by spiking solvent carriers, such as the loss of hydrophobic compounds through binding with plastic surfaces [17]. In TEST II, C. elegans was exposed to a maximum water-soluble concentration for only for 3 hours, because behavioral abnormalities were observed after 6 hours when C. elegans was exposed to 400 μg/L 9-MA. This partly supports that toxic effects of alkyl-Ant could be seen at lower levels than observed in earlier studies via oxidative stress.